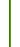В ходе обсуждения статей о потомках шумеров стало очевидно, что эта тема, как и неотделимый от нее вопрос о происхождении шумеров как этноса, требует знания материала по ДНК не только современных жителей Ирака и стран Персидского залива, но и примыкающих к нему регионов. Без детальной информации о распределении и датировках различных генеалогических линий у народов, населяющих Средиземноморье, Ближний Восток, Переднюю, Центральную и Южную Азию нет возможности обосновать, какая из возможных гипотез о миграциях предков шумеров (и части современных арабов, соответственно) описывает события наиболее близко к истине. Только при столь масштабном анализе можно выявить направления и вероятные датировки миграций в интересующем нас регионе и согласовать эти предположения с материалом из других дисциплин, в первую очередь археологии.

Как уже говорилось в первой части статьи, существующие полевые выборки Y-хромосомных гаплотипов, при всей их статистической значимости, недостаточно информативны для поставленной задачи в силу того, что в них собраны, в лучшем случае, 17-маркерные гаплотипы в формате YFiler, причем зачастую без отнесения по снипам. При столь низком разрешении ни о каком выделении и анализе специфических линий, подобном тому, что был проделан во второй части статьи, не может быть речи. Они или окажутся неотличимыми от других ветвей той же гаплогруппы/субклада, или, что еще хуже, одна и та же ветвь будет разделена на фрагменты, родство которых доказать крайне сложно. Особенно это актуально для линий, зародившихся в интересующий нас промежуток времени, т.е. более 5000 лет назад.
По этой причине адекватному анализу поддаются только выборки протяженных гаплотипов (37 маркеров и более) с проектов FTDNA, даже при столь очевидном их недостатке, как крайне неравномерная представленность в них разных стран и этнических групп. Например, участники из Саудовской Аравии (29 млн. населения) исчисляются тысячами (не менее 3000), а Эфиопию (более 90 млн.) представляет от силы 10-15 человек. Поскольку расчетный аппарат ДНК-генеалогии менее чувствителен к статистике гаплогрупп по регионам, чем методы популяционной генетики, дефицит информации по таким народам, как курды, азербайджанцы, дагестанцы или персы не должен радикально повлиять на общие выводы из анализа. С другой стороны, очевидно, что для серьезного анализа требуется выборка как можно большего размера, чтобы получить репрезентативную статистику по минорным линиям, без которой сложно подтвердить или опровергнуть предлагаемые трактовки.
Такая выборка в количестве 9393 гаплотипов была составлена по данным с 66 географических проектов FTDNA, от Пиренейского полуострова на западе до Индостана на востоке. Статистика по регионам представлена в графическом виде ниже.
Поскольку очевидно, что генеалогические линии, как правило, сформировались задолго до установления нынешних политических и этнических границ, данные по отдельным странам и народам сгруппированы по регионам, более релевантным поставленной задаче. С запада на восток это Иберия (Испания, Португалия и страны Латинской Америки), Италия (включая Сицилию, Сардинию и Мальту), Магриб (включая арабов и берберов стран Сахеля), Балканы (включая острова Восточного Средиземноморья), долина Нила, Анатолия (Турция, Армения, северо-запад Ирана, а также ассирийцы и армяне из Ливана и Ирака), Левант, Персидский залив, Южная Аравия, Закавказье (Грузия и Азербайджан), Северный Кавказ (осетины, карачаевцы, балкарцы, чеченцы и ингуши, остальные народы представлены единичными гаплотипами) и страны Средней и Южной Азии (включая этнических персов Ирана). Евреи ашкенази не включены в список, поскольку они сформировались как этническая группа за пределами рассматриваемого здесь региона. В раздел «Прочие» (Other) помещены гаплогруппы, доля которых слишком мала для надежного анализа, либо те, что специфичны для других регионов. Например, это африканские гаплогруппы А и В, что найдены у арабов, гаплогруппа Н, распространенная в Индии, или субклад Q1a2a1 (M54) у жителей Латинской Америки – потомков ее коренного населения.
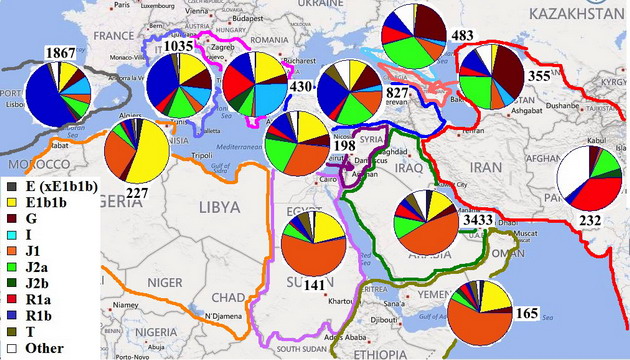
Напомню еще раз, по каким признакам велся поиск линий, что могут восходить к шумерам. Они должны быть достаточно специфическими для региона Персидского залива, представлять там статистически значимую долю населения и сходиться к предку, жившему между 5700 и 5200 годами до настоящего времени. В предыдущей статье наиболее вероятными кандидатами в такие линии были названы ветви, принадлежавшие субкладам J2a1* (PF5197) и J2a1h2* (F3133). Для удобства, они выделены на упрощенном дереве гаплогруппы J2 фиолетовым и зеленым цветом, соответственно.
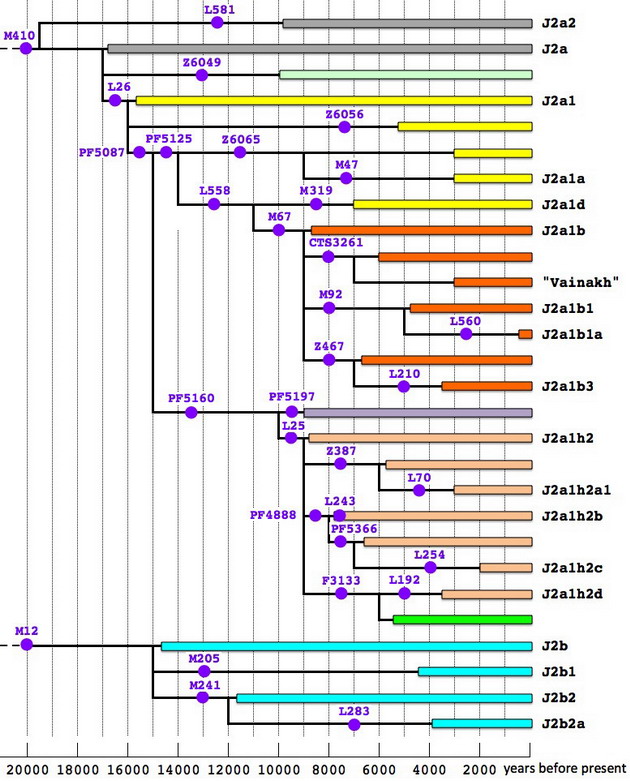
А вот как выглядит в графической форме распределение разных ветвей гаплогруппы J2 по всем 12 регионам. Высота столбика на диаграмме пропорциональна общей доле J2 в соответствующей выборке, а цвет отмечает группы ветвей, выделенных таким же образом на дереве.
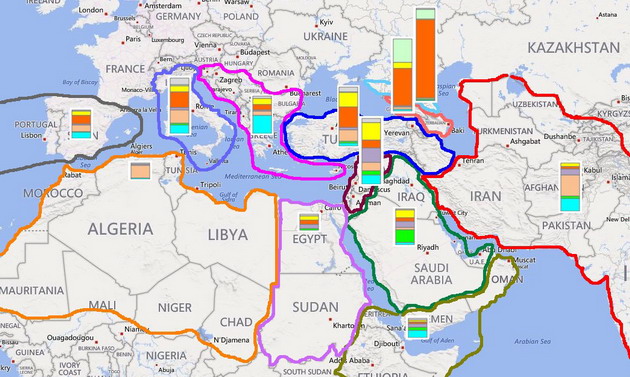
Обращает на себя внимание то, что, несмотря на древность большинства ветвей гаплогруппы J2, они весьма выраженно расходятся по своей географии. Так, субклад J2a1b (M67) незначительно представлен к югу и востоку от Анатолии, но доминирует у народов Закавказья и Северного Кавказа, а также является одним из основных у итальянцев и жителей Пиренейского полуострова. Видимо, пути носителей этой старой (около 9000 лет до общего предка) ветви пролегали от Анатолии (вероятной прародины?) в западном и северо-восточном направлениях, но почти не затрагивали южную часть Плодородного Полумесяца, где в эпоху раннего неолита, видимо, доминировали «афразийцы» из различных ветвей гаплогруппы Е.
Что касается уже обсуждавшейся ранее «зеленой» ветви F3133, то данные с более масштабной выборки подтвердили сделанные тогда выводы. По всем трем признакам она соответствует генеалогической линии, зародившейся в среде жителей первых поселений в низовьях Тигра и Евфрата времен Среднего Урукского периода, т.е. шумеров. Другая линия, специфическая для арабов Персидского залива, PF5197, является одной из самых старых в регионе (около 9000 лет до предка) и показывает пик своего распространения в Леванте. На упрощенной схеме детальная филогения этой ветви опущена, но дочерние ветви со своей специфической географией можно отметить на дереве, построенном по списку 37-маркерных гаплотипов, подтвержденных как PF5197.
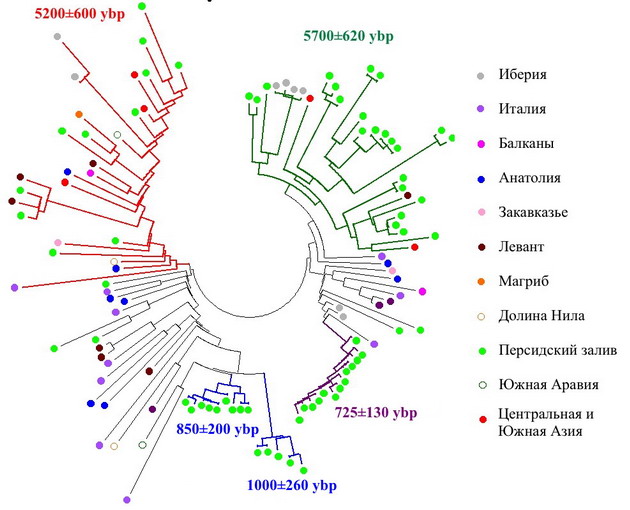
Ветвь, расположенная в верхней правой части диаграммы и, возможно, имеющая собственный снип PF5174, почти целиком состоит из арабов Персидского залива, а время жизни ее предка попадает в тот же самый интервал, что для ветви F3133. Еще 3 арабские ветви с недавними общими предками, но сходящиеся ко времени около 6200 лет назад подтверждают, что представители субклада PF5174 населяют берега Персидского залива с древних времен. Исходя из географии PF5174 и родственного ему субклада J2a1h2 (L25), можно ориентировочно оценить, где около 10000 лет назад жил их общий предок и откуда, по-видимому, начали свой путь люди, переселившиеся в IV тысячелетии до н.э. на берега Тигра и Евфрата. Поскольку ветви PF5197 и L25 практически отсутствуют в Кавказском регионе, можно предположить, что их общий предок (PF5160) жил в другом месте, нежели его современник – предок специфического для Кавказа субклада J2a1b (М67).
С другой стороны, оно не должно пересекаться с регионом, где во времена неолита преобладала гаплогруппа Е, поскольку ее представители отсутствуют в Средней и Южной Азии, где субклад J2a1h2 (L25) представлен несколькими ветвями. Методом исключения остается северная часть Леванта или примыкающие к нему регионы Малой Азии, включая острова Восточного Средиземноморья. С древнейших времен это был один из центров мореплавания, благодаря которому, очевидно, представители различных ветвей PF5197 и L25 заселили все Средиземноморье. Навыки мореплавания наверняка пригодились и при расселении в восточном направлении – вдоль побережья Персидского залива к долине Инда, где мы наблюдаем высокую долю субклада J2a1h2 (L25). Как и в ситуации с шумерами, налицо следы миграции со стороны Восточного Средиземноморья, но по Индии и Пакистану пока не так много данных, чтобы выявить основные ветви, рассчитать их датировки и оценить возможный вклад ассоциирующихся с ними народов в становление древнеиндийских цивилизаций. Вопрос о происхождении и этнической принадлежности людей, создавших первые городские цивилизации долины Инда до сих пор остается открытым, и без данных ДНК-генеалогии ответ на него вряд ли возможен.
Завершая разбор гаплогруппы J2а, хотел бы отметить, что в последние несколько лет ее стали ассоциировать с распространением индоевропейских языков. Более того, использовать такое отнесение как один из аргументов в поддержку анатолийской гипотезы о происхождении индоевропейской семьи, в чем подписался и один из ее авторов – К. Ренфрю. Как мы видим из анализа, датировки, предполагаемые места зарождения и некоторые миграционные пути основных ветвей J2a, хоть и с натяжками, но согласуются с этой гипотезой. Однако при более глубоком анализе становится ясно, что их носители в то время говорили на языках, не имевших отношения к индоевропейским. Возможно, шумерский является единственным реликтом той исчезнувшей языковой семьи, что дожил до письменной фиксации и был расшифрован. Другим повезло меньше, и от них, в лучшем случае, остались не поддающиеся прочтению надписи, что находят по всему Восточному Средиземноморью.
А что же эрбины? Каков их вклад в шумерскую цивилизацию? Очевидно, новую информацию можно получить по той же схеме, что использовалась выше для гаплогруппы J2. Вот как выглядят упрощенное дерево гаплогруппы R1b и карта ее распространения, соответственно.
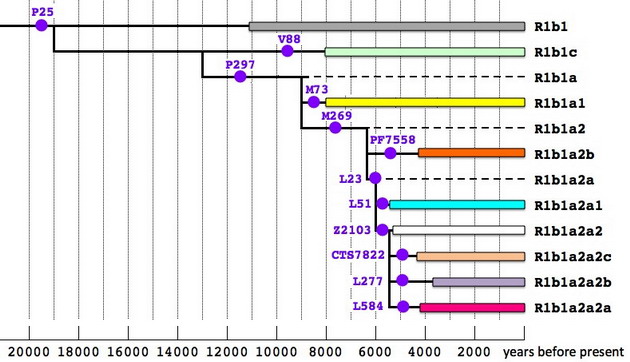
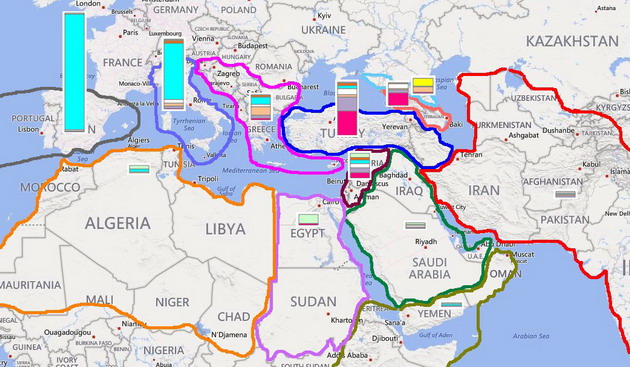
Принципы обозначений те же самые, что и для J2. На дереве пунктиром обозначены парагруппы, для которых пока не найдено подтвержденных носителей, а белым цветом – как гаплотипы из родительской ветви Z2103 (их меньшинство) так и те, что не удается пока отнести к какой-либо из ее трех принципиальных подветвей из-за отсутствия данных по снипам.
Как и в предыдущем исследовании, выраженный пик «восточных» ветвей гаплогруппы R1b приходится на Анатолию, которая в проектах FTDNA представлена в основном армянами, тогда как в регионах Персидского залива, Южной Аравии, Средней и Южной Азии эта гаплогруппа относится к минорным. По-видимому, пути эрбинов в Европу проходили в стороне от низовьев Тигра и Евфрата, и их вклад в этногенез шумеров был невелик. Корреляции (по крайней мере) ветвей L277 и L584 с потомками хурритов и урартов находятся в большем согласии с данными письменной истории и лингвистики. С другой стороны, собранные в этом исследовании данные позволяют отодвинуть время появления носителей R1b с 6500-6000 лет назад на более раннюю дату. В пользу того, что эрбины начали свой путь из Центральной Азии еще до времени расхождения основных ветвей субклада R1b1a2 (по которому и делалась оценка), говорит тот факт, что представитель парагруппы R1b1* (P25) был найден в Испании, в захоронении времен культуры импрессо (кардиальной керамики), с калиброванной радиоуглеродной датировкой 7200-7080 лет назад. Эту культуру раннего неолита, отмеченную на северном побережье Средиземного моря, от Адриатики до Каталонии, связывают с миграциями из Леванта или Малой Азии, начавшимися не менее 8000 лет назад. Очевидно, эрбины появились на Ближнем Востоке, по меньшей мере, не позже этой даты.
Ветви соответствующего «возраста» на дереве гаплогруппы R1b имеются. Наряду с парагруппой R1b1* (P25), что рассеяна с чрезвычайно низкой частотой от Испании до Индии и Узбекистана, их можно считать реликтами той давней миграции. Это субклад R1b1c (V88), специфический для Ближнего Востока и Африки, и R1b1a1 (M73), что встречается в основном у жителей евразийских степей, но одна из ветвей которого найдена у итальянцев и испанцев. В выборке из долины Нила (Египет и Судан) ветвь V88 представлена 6 из 7 гаплотипов, относящихся к R1b, что вполне ожидаемо для региона, расположенного на пути от Леванта к озеру Чад. Лингвисты находят специфический «северокавказский» субстрат в чадских языках афразийской макросемьи, но никто не смог внятно объяснить, каким образом он там появился. Из данных ДНК этот вывод очевиден, если принять во внимание, что субклад V88 достигает пика среди народов, говорящих на языках чадской группы. Следовательно, субстрат они унаследовали от языка народа, родственного другим ветвям эрбинов. Из схем также можно заключить, что эрбины, видимо, принимали участие в этногенезе древних египтян, но еще в додинастические времена, насколько можно судить по датировкам субклада V88.
Когда же эти древние ветви R1b появились на Ближнем Востоке? Есть ли какие-нибудь независимые указания на их путь из Центральной Азии, или правы те, чье мнение изложено в статье в Википедии, и родина R1b – Ближний Восток? В одном из прежних комментариев я указывал на одно из косвенных подтверждений того, что какая-то группа людей пришла на Ближний Восток со стороны Центральной Азии между 8000 и 9000 годами назад. Это появление керамики, которую изобрели около 19000 лет назад в Восточной Азии. Однако можно резонно возразить, что изобретения подобного рода делались независимо в разное время и в разных частях Земли, а потому древним жителям Ближнего Востока вовсе не нужно было перенимать эту технологию у тех, кто ей владел задолго до того. Ввиду очень скудного материала по самым ранним образцам керамики однозначного ответа пока нет.
Но есть и другой пример, более объективный. Это просо (Panicum miliaceum L.), процесс одомашнивания которого уже не одно десятилетие остается одной из нерешенных загадок в эволюционной биологии. Об этой проблеме в контексте древних связей между Востоком и Западом можно в общих чертах узнать из лекции К. Ренфрю (на английском, с 22 по 25 минуты) на конференции 2011 года, посвященной Великому Шелковому пути. Дикорастущие формы этого злака неизвестны, а самые ранние находки одомашненного проса датируются около 10000 лет назад на севере Китая и, самое позднее, 7000 лет назад в Закавказье. Чтобы разрешить загадку раннего появления этой сельскохозяйственной культуры в столь удаленных друг от друга местах, была предложена версия о независимом одомашнивании. Недавно ее проверили генетики, исследовавшие ДНК проса из 98 мест в Евразии (H.V. Hunt et al. Genetic diversity and phylogeography of broomcorn millet (Panicum miliaceum L.) across Eurasia. Mol Ecol. 2011 Nov; 20(22): 4756–4771). Авторы проанализировали 25-маркерные наборы аллелей из всех образцов, но так и не пришли к выводу, было ли просо одомашнено один раз в Китае и затем его культура была принесена в западную часть Евразии, или из Китая в Переднюю Азию попала одна из дикорастущих форм, и ее одомашнили независимо. Вот как выглядят результаты разложения по компонентам, где каждый из рассчитанных генетических пулов выделен цветом
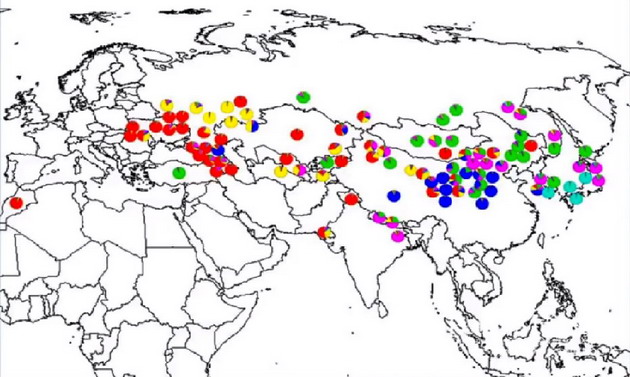
Осторожность авторов в выводах понятна, потому что довольно низкое разрешение геномных данных оставляет простор для интерпретаций (сравните 25 маркеров с миллионами нуклеотидов при анализе генома человека), но очевидно, что существовал один центр, из которого пошло все генетическое разнообразие проса. Это район современных китайских провинций Ганьсу, Шаньси и Шэньси, что согласуется и с палеоботаническими данными. Достиг предок европейских разновидностей проса (красные и желтые сектора) своего нынешнего ареала в диком или одомашненном состоянии, данные ДНК сами по себе сказать не могут, что, собственно, и констатировали авторы работы. Однако если к ним добавить отсутствие дикорастущих форм проса (вероятность сохранения которых была бы выше в случае двух центров) и данные ДНК-генеалогии о гаплогруппах, что существовали в степной зоне Евразии в начале послеледниковой эпохи, то чаша весов склонится в пользу миграции людей, принесших с собой эту очень неприхотливую и засухоустойчивую культуру из степей Центральной Азии.
Методом исключения основной генеалогической линией этих людей оказывается гаплогруппа R (к которой принадлежал, по крайней, один житель Прибайкалья 24 тысячи лет назад), с Q (Q1b?) в качестве минорной линии. Датировки первых находок проса (а также керамической посуды, что необходима для приготовления каши из мелких и непригодных для выпечки хлеба просяных зерен) на западе Евразии и времена жизни предков самых старых ветвей гаплогруппы R1b совпадают в пределах погрешности, тем самым давая дополнительный аргумент в пользу миграции эрбинов из Центральной Азии. Однако к возникновению шумерской цивилизации они имели очень опосредованное отношение, как можно заключить по сумме данных.
Наконец, коротко остановимся на еще одной генеалогической линии, что в древние времена прошла «великим просяным путем» и также внесла свой вклад в этногенез народов Ближнего Востока. Это гаплогруппа R1a, упрощенное дерево и карта распределения которой приведены ниже.
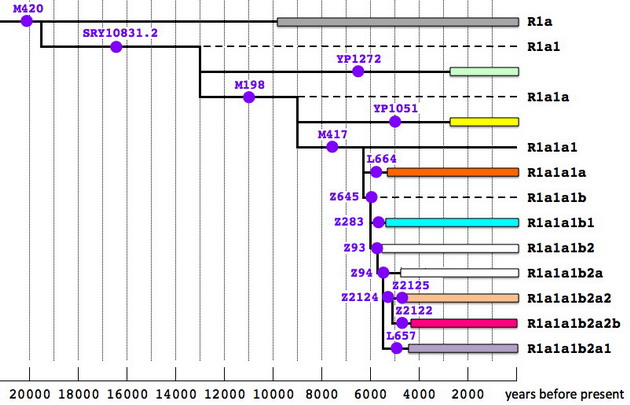
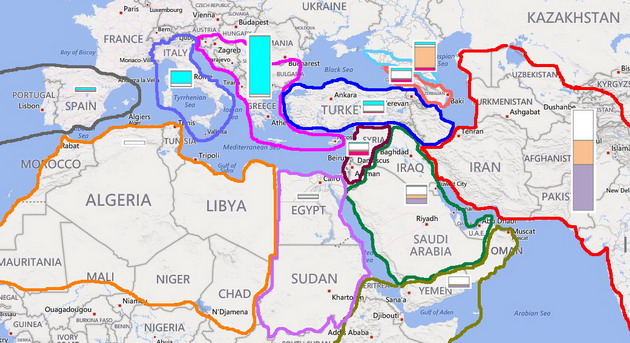
Обозначения на схемах следуют тем же самым правилам, что и в предыдущих случаях. Нельзя не обратить внимание, что деревья гаплогрупп R1a и R1b похожи по структуре и датировкам ветвлений, как близнецы. Однако есть и существенное различие. Реликтовые ветви R1a найдены пока почти исключительно в Европе, а их аналоги из R1b тяготеют к Азии и Ближнему Востоку. Очевидно, это указывает на раннее географическое расхождение двух родственных ветвей, пути которых вновь пересеклись лишь в эпоху энеолита.
Карта распределения субкладов гаплогруппы R1a позволяет с достаточно высокой вероятностью выявить миграционные потоки, приведшие эту гаплогруппу на Ближний Восток. Первый, самый массовый – от евразийских степей к Индии, Иранскому нагорью и далее к Персидскому заливу, – характеризуют субклады L657 и Z2125 (родительский к «туранскому» Z2123). Второй маршрут – от причерноморских степей через Кавказ в Закавказье, Анатолию и Левант, – можно отследить по субкладу Z2122, но для более детальной информации необходимы данные по участникам ДНК-проектов из Грузии и Турции, которые в основной своей массе не анализировали снипы. Возможно, был еще один путь, самый ранний – со стороны Балканского полуострова через черноморские проливы. Его маркируют гаплотипы из субклада Z283, среди которых заметную долю занимают представители родительской ветви, а в качестве минорных линий отмечены родительская ветвь Z93 и реликтовая R1a* (M420). Однако с равной вероятностью они могли попасть в Анатолию и через Кавказ, по которому пока данных явно недостаточно.
Любопытно, что «анатолийские» ветви R1b-L277 и R1b-L284 дают примерно те же датировки, что и специфическая для этого же региона ветвь R1a-Z2122. Вполне вероятен вариант, что хурриты (R1b?) и митаннийские арии (R1a?) появились в Анатолии в одно и то же время в составе одного и того же союза племен, имевших сходную материальную культуру, а потому неразличимых по археологическим находкам. Во всяком случае, пестрый этнический состав созданного ими государства Митанни был установлен в основном по эпиграфическим данным, а не по керамике или предметам обихода. Подобные коалиции разных народов, объединявшихся во время миграции – далеко не редкость в истории. Классические примеры: кимвры и тевтоны, аланы и вандалы, а также всевозможные тюрко-монголо-маньчжурские конфедерации, вторгавшиеся в Китай. Однако это уже отдельная тема, не имеющая прямого отношения к шумерам, в этногенезе которых доля гаплогруппы R1a была крайне незначительной, если не нулевой.
В заключение можно сказать, что собранный в этом исследовании материал позволил с большей, чем ранее, надежностью реконструировать детали истории Месопотамии времен зарождения шумерских городов-государств. Несомненна их связь с более ранними культурами, существовавшими в Восточном Средиземноморье. Хотелось бы надеяться, что данные ДНК-генеалогии дадут подсказку археологами и историкам, где искать эти культуры и/или как интерпретировать уже имеющиеся находки. Круг вопросов, рассмотренных в этой статье, вышел за пределы заявленной в ней темы. Надеюсь, читатели воспримут это не как уход от предмета обсуждения, а как прелюдию к последующему анализу материала в более широкой перспективе.
Игорь Рожанский,
кандидат химических наук
Источник |